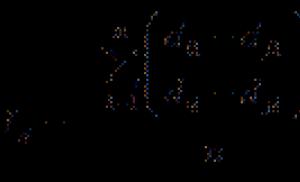Генетическое разнообразие народов. Этногеномика и генетическая история народов восточной европы Генетический метод расселения народов недостатки
| Содержание |
|---|
| Генетическое разнообразие народов |
| Происхождение и расселение человека |
| Адаптация к различным условиям обитания |
| Устойчивость к инфекционным заболеваниям |
| Развитие цивилизации и генетические изменения |
| Заключение |
| Литература |
| Все страницы |
Страница 2 из 7
Происхождение и расселение человека
Ранее историю появления вида Homo sapiens на Земле реконструировали на основе палеонтологических, археологических и антропологических данных. В последние десятилетия появление молекулярно-генетических методов и исследования генетического разнообразия народов позволили уточнить многие вопросы, связанные с происхождением и расселением людей современного анатомического типа.
Молекулярно-генетические методы, используемые для восстановления демографической истории, сходны с лингвистической реконструкцией праязыка. Время, когда два родственных языка разделились (т.е. когда исчез их общий предковый праязык), оценивают по количеству различающихся слов, появившихся за период раздельного существования этих языков. Аналогично возраст предковой популяции, общей для двух современных народов, рассчитывают по количеству мутаций, накопившихся в ДНК их представителей. Чем больше различий в ДНК, тем больше времени прошло с момента разделения популяций. Так как скорость накопления мутаций в ДНК известна, по числу мутаций, отличающих две популяции, можно определить дату их расхождения (если предположить, что после разделения они больше не встречались и не смешивались).
Для датировки этого события используют нейтральные мутации, которые не влияют на жизнеспособность индивида и не подвержены действию естественного отбора. Они найдены во всех участках генома человека, но наиболее часто используют мутации в ДНК, содержащейся в клеточных органеллах - митохондриях. В оплодотворенной яйцеклетке присутствует только материнская митохондриальная ДНК (мтДНК), поскольку спермий свои митохондрии яйцеклетке не передает. Для филогенетических исследований мтДНК имеет особые преимущества. Во-первых, она не подвергается рекомбинации, как аутосомные гены, что значительно упрощает анализ родословных. Во-вторых, в клетке она содержится в количестве нескольких сот копий и гораздо лучше сохраняется в биологических образцах.
Первым использовал мтДНК для реконструкции истории человечества американский генетик Алан Уилсон в 1985 г. Он изучил образцы мтДНК, полученные из крови людей из всех частей света, и на основе выявленных между ними различий построил филогенетическое древо человечества. Оказалось, что все современные мтДНК могли произойти от мтДНК общей праматери, жившей в Африке. Обладательницу предковой мтДНК тут же окрестили “митохондриальной Евой”, что породило неверные толкования - будто все человечество произошло от одной-единственной женщины. На самом деле у “Евы” было несколько тысяч соплеменниц, просто их мтДНК до наших времен не дошли. Однако все они, без сомнения, оставили свой след: от них мы унаследовали генетический материал хромосом. Характер наследования в данном случае можно сравнить с семейным имуществом: деньги и земли человек может получить от всех предков, а фамилию - только от одного из них. Генетическим аналогом фамилии, передаваемой по женской линии, служит мтДНК, а по мужской - Y-хромосома, передаваемая от отца к сыну.
Изучение мтДНК и ДНК Y-хромосомы подтвердили африканское происхождение человека, позволили установить пути и даты его миграции на основе распространения различных мутаций у народов мира. По современным оценкам, вид H.sapiens появился в Африке более 100 тыс. лет назад, затем расселился в Азии, Океании и Европе. Позже всего была заселена Америка.
Вероятно, исходная предковая популяция H.sapiens состояла из небольших групп, ведущих жизнь охотников-собирателей. Мигрируя, люди несли с собой свои традиции, культуру и свои гены. Возможно, они также обладали и праязыком. Пока лингвистические реконструкции происхождения языков мира ограничены 15-30 тыс. лет, и существование общего праязыка только предполагается. И хотя гены не определяют ни язык, ни культуру, в некоторых случаях генетическое родство народов совпадает и с близостью их языков и культурных традиций. Но есть и противоположные примеры, когда народы меняли язык и перенимали традиции своих соседей. Такая смена происходила чаще в районах контактов различных волн миграций или же в результате социально-политических изменений или завоеваний.
Конечно, в истории человечества популяции не только разделялись, но и смешивались. На примере линий мтДНК результаты такого смешения можно наблюдать у народов Волго-Уральского региона. Здесь столкнулись две волны расселения, европейская и азиатская. В каждой из них к моменту встречи на Урале в мтДНК успели накопиться десятки мутаций. У народов Западной Европы азиатские линии мтДНК практически отсутствуют. В Восточной Европе они встречаются редко: у словаков с частотой 1%, у чехов, поляков и у русских Центральной России - 2%. По мере приближения к Уралу частота их возрастает: у чувашей - 10%, у татар - 15%, у разных групп башкир - 65-90%. Закономерно, что у русских Волго-Уральского региона количество азиатских линий больше (10%), чем в Центральной России.
научн. сотр. лаборатории анализа генома Ин-та
общей генетики
им. Н.И. Вавилова РАН
Генетическое разнообразие народов
 Люди, живущие в разных
концах Земли, отличаются по многим признакам:
языковой принадлежности, культурным традициям,
внешности, генетическим особенностям.
Генетические характеристики народов зависят от
их истории и образа жизни. Различия между ними
возникают в изолированных популяциях, не
обменивающихся потоками генов (т.е. не
смешивающихся из-за географических,
лингвистических или религиозных барьеров), за
счет случайных изменений частот аллелей и
процессов позитивного и негативного
естественного отбора.
Люди, живущие в разных
концах Земли, отличаются по многим признакам:
языковой принадлежности, культурным традициям,
внешности, генетическим особенностям.
Генетические характеристики народов зависят от
их истории и образа жизни. Различия между ними
возникают в изолированных популяциях, не
обменивающихся потоками генов (т.е. не
смешивающихся из-за географических,
лингвистических или религиозных барьеров), за
счет случайных изменений частот аллелей и
процессов позитивного и негативного
естественного отбора.
Случайное изменение частот аллелей в популяции называется генетическим дрейфом . Различия этих частот без действия каких-либо дополнительных факторов обычно невелики. При сокращении численности или отселении небольшой группы, дающей начало новой популяции, частоты аллелей могут сильно колебаться. В новой популяции они будут зависеть от генофонда основавшей ее группы (так называемый эффект основателя – все носители мутации получают ее от общего предка, у которого она возникла). С этим эффектом связывают повышенную частоту болезнетворных мутаций в некоторых этнических группах. Например, у японцев один из видов врожденной глухоты вызывается мутацией, возникшей однократно в прошлом и не встречающейся в других районах мира. У белых австралийцев глаукома связана с мутацией, завезенной переселенцами из Европы. У исландцев найдена мутация, повышающая риск развития рака и восходящая к общему прародителю. Аналогичная ситуация обнаружена у жителей о. Сардиния, но у них мутация другая, отличная от исландской. Среди русских, живущих в Башкортостане, из нескольких сотен мутаций, приводящих к фенилкетонурии, встречается преимущественно одна, что связывают с переселением в этот регион относительно небольшой группы русских, обладавших ею. Эффект основателя – одно из возможных объяснений отсутствия у американских индейцев разнообразия по группам крови AB0: у них преобладает группа 0 (первая), частота ее более 90%, а во многих популяциях – 100%. Так как Америка заселялась небольшими группами, пришедшими из Азии через перешеек, соединявший эти материки десятки тысяч лет назад, возможно, что в популяции, давшей начало коренному населению Нового Света, другие группы крови отсутствовали.
Слабовредные мутации могут долго поддерживаться в популяции, а вредные, значительно снижающие приспособленность индивида, отсеиваются отбором. Показано, что болезнетворные мутации, вызывающие тяжелые формы наследственных заболеваний, обычно эволюционно молоды. Давно возникшие мутации, длительное время сохраняющиеся в популяции, связаны с более легкими формами болезни.
Адаптация к условиям обитания фиксируется в ходе отбора благодаря случайно возникшим новым аллелям, повышающим приспособленность к данным условиям, или за счет изменения частот давно существующих аллелей. Разные аллели обусловливают варианты фенотипа, например цвета кожи или уровня холестерина крови. Частота аллеля, обеспечивающего адаптивный фенотип (например, темная кожа в зонах с интенсивным солнечным облучением), возрастает, поскольку его носители более жизнеспособны в данных условиях.
Адаптация к различным климатическим зонам проявляется как вариация частот аллелей комплекса генов, географическое распределение которых соответствует климатическим зонам. Однако наиболее заметный след в глобальном распределении генетических изменений оставили миграции народов, связанные с расселением от африканской прародины.
Происхождение и расселение человека
Ранее историю появления вида Homo sapiens на Земле реконструировали на основе палеонтологических, археологических и антропологических данных. В последние десятилетия появление молекулярно-генетических методов и исследования генетического разнообразия народов позволили уточнить многие вопросы, связанные с происхождением и расселением людей современного анатомического типа.
Молекулярно-генетические методы, используемые для восстановления демографической истории, сходны с лингвистической реконструкцией праязыка. Время, когда два родственных языка разделились (т.е. когда исчез их общий предковый праязык), оценивают по количеству различающихся слов, появившихся за период раздельного существования этих языков. Аналогично возраст предковой популяции, общей для двух современных народов, рассчитывают по количеству мутаций, накопившихся в ДНК их представителей. Чем больше различий в ДНК, тем больше времени прошло с момента разделения популяций. Так как скорость накопления мутаций в ДНК известна, по числу мутаций, отличающих две популяции, можно определить дату их расхождения (если предположить, что после разделения они больше не встречались и не смешивались).
Для датировки этого события используют нейтральные мутации, которые не влияют на жизнеспособность индивида и не подвержены действию естественного отбора. Они найдены во всех участках генома человека, но наиболее часто используют мутации в ДНК, содержащейся в клеточных органеллах – митохондриях. В оплодотворенной яйцеклетке присутствует только материнская митохондриальная ДНК (мтДНК), поскольку спермий свои митохондрии яйцеклетке не передает. Для филогенетических исследований мтДНК имеет особые преимущества. Во-первых, она не подвергается рекомбинации, как аутосомные гены, что значительно упрощает анализ родословных. Во-вторых, в клетке она содержится в количестве нескольких сотен копий и гораздо лучше сохраняется в биологических образцах.
Первым использовал мтДНК для реконструкции истории человечества американский генетик Алан Уилсон в 1985 г. Он изучил образцы мтДНК, полученные из крови людей из всех частей света, и на основе выявленных между ними различий построил филогенетическое древо человечества. Оказалось, что все современные мтДНК могли произойти от мтДНК общей праматери, жившей в Африке. Обладательницу предковой мтДНК тут же окрестили «митохондриальной Евой», что породило неверные толкования – будто все человечество произошло от одной-единственной женщины. На самом деле у «Евы» было несколько тысяч соплеменниц, просто их мтДНК до наших времен не дошли. Однако все они, без сомнения, оставили свой след: от них мы унаследовали генетический материал хромосом. Характер наследования в данном случае можно сравнить с семейным имуществом: деньги и земли человек может получить от всех предков, а фамилию – только от одного из них. Генетическим аналогом фамилии, передаваемой по женской линии, служит мтДНК, а по мужской – Y-хромосома, передаваемая от отца к сыну.
Изучение мтДНК и ДНК Y-хромосомы подтвердили африканское происхождение человека, позволили установить пути и даты его миграции на основе распространения различных мутаций у народов мира. По современным оценкам, вид H.sapiens появился в Африке более 100 тыс. лет назад, затем расселился в Азии, Океании и Европе. Позже всего была заселена Америка.
Вероятно, исходная предковая популяция H.sapiens состояла из небольших групп, ведущих жизнь охотников-собирателей. Мигрируя, люди несли с собой свои традиции, культуру и свои гены. Возможно, они также обладали и праязыком. Пока лингвистические реконструкции происхождения языков мира ограничены периодом 15–30 тыс. лет, и существование общего праязыка только предполагается. И хотя гены не определяют ни язык, ни культуру, в некоторых случаях генетическое родство народов совпадает и с близостью их языков и культурных традиций. Но есть и противоположные примеры, когда народы меняли язык и перенимали традиции своих соседей. Такая смена происходила чаще в районах контактов различных волн миграций или же в результате социально-политических изменений или завоеваний.
Конечно, в истории человечества популяции не только разделялись, но и смешивались. На примере линий мтДНК результаты такого смешения можно наблюдать у народов Волго-Уральского региона. Здесь столкнулись две волны расселения – европейская и азиатская. В каждой из них к моменту встречи на Урале в мтДНК успели накопиться десятки мутаций. У народов Западной Европы азиатские линии мтДНК практически отсутствуют. В Восточной Европе они встречаются редко: у словаков – с частотой 1%, у чехов, поляков и русских Центральной России – 2%. По мере приближения к Уралу частота их возрастает: у чувашей – 10%, у татар – 15%, у разных групп башкир – 65–90%. Закономерно, что у русских Волго-Уральского региона количество азиатских линий больше (10%), чем в Центральной России.
К изменениям условий среды (температуры, влажности, интенсивности солнечного облучения) человек приспосабливается за счет физиологических реакций (потоотделения, загара и т.п.). Однако в популяциях, проживающих долгое время в определенных климатических условиях, адаптации к ним накапливаются на генетическом уровне. Они меняют внешние признаки, сдвигают границы физиологических реакций (например, скорость сужения сосудов конечностей при охлаждении), «подстраивают» биохимические параметры (такие, как уровень холестерина в крови) к оптимальным для данных условий.
Климат. Один из наиболее известных расовых признаков – цвет кожи, пигментация которой у человека задана генетически. Пигментация защищает от повреждающего действия солнечного облучения, но не должна препятствовать получению минимальной дозы облучения, необходимой для образования витамина D, предотвращающего рахит. В северных широтах, где низкая интенсивность облучения, у людей кожа более светлая, а в экваториальной зоне – самая темная. Однако у обитателей затененных тропических лесов кожа светлее, чем можно было бы ожидать на данной широте, а у некоторых северных народов (чукчей, эскимосов), напротив, она относительно сильно пигментирована. В последнем случае это объясняется либо поступлением витамина D с пищей (рыбой и печенью морских животных), либо недавней в эволюционном масштабе миграцией северных групп из более низких широт.

Таким образом, интенсивность ультрафиолетового излучения действует как фактор отбора, приводя к географическим вариациям в цвете кожи. Светлая кожа – эволюционно более поздний признак и возникла за счет мутаций в нескольких генах, регулирующих выработку кожного пигмента меланина (ген рецептора меланинокортина MC1R и другие). Способность загорать также детерминирована генетически. Ею отличаются жители регионов с сильными сезонными колебаниями интенсивности солнечного излучения.
Известны связанные с климатическими условиями различия в строении тела. Это адаптации к холодному или теплому климату. Так, короткие конечности у жителей арктических областей (чукчей, эскимосов) уменьшают отношение поверхности тела к его массе и тем самым сокращают теплоотдачу. Обитатели жарких сухих регионов, например африканские масаи, напротив, отличаются длинными конечностями. У жителей влажного климата более широкие и плоские носы, а в сухом и более холодном климате носы длиннее, что способствует согреванию и увлажнению вдыхаемого воздуха.
Повышенное содержание гемоглобина в крови и усиление легочного кровотока служат приспособлением к высокогорным условиям. Такие особенности свойственны аборигенам Памира, Тибета и Анд. Все эти признаки определяются генетически, но степень их проявления зависит от условий развития в детстве: например, у андских индейцев, выросших на уровне моря, а затем переселившихся в высокогорные районы они менее выражены.
Типы питания. Некоторые генетические изменения связаны с разными типами питания. Среди них наиболее известна непереносимость молочного сахара (лактозы) – гиполактазия. У детенышей всех млекопитающих для усвоения лактозы вырабатывается фермент лактаза. По окончании вскармливания она исчезает из кишечного тракта детеныша. Отсутствие фермента у взрослых – исходный, предковый признак для человека.
Во многих азиатских и африканских странах, где взрослые традиционно не пьют молоко, после пятилетнего возраста лактаза не синтезируется, и потому употребление молока приводит к расстройству пищеварения. Однако большинство взрослых европейцев могут без вреда для здоровья пить молоко: синтез лактазы у них не прекращается из-за мутации в участке ДНК, регулирующем образование фермента. Эта мутация распространилась после появления молочного скотоводства 9–10 тыс. лет назад и встречается преимущественно у европейских народов. Более 90% шведов и датчан способны усваивать молоко, и лишь небольшая часть населения Скандинавии отличается гиполактазией. В России частота гиполактазии составляет около 30% для русских и более 60–80% для коренных народов Сибири и Дальнего Востока. Народы, у которых гиполактазия сочетается с молочным скотоводством, традиционно используют не сырое молоко, а кисломолочные продукты, в которых молочный сахар уже расщеплен бактериями.
Отсутствие сведений о генетических особенностях народов порой приводит к тому, что при гиполактазии людям, реагирующим на молоко расстройством пищеварения, которое принимают за кишечные инфекции, вместо необходимого изменения диеты предписывают лечение антибиотиками, ведущее к дисбактериозу.
Кроме употребления молока еще один фактор мог влиять на сохранение у взрослых синтеза лактазы. В присутствии лактазы молочный сахар способствует усвоению кальция, выполняя те же функции, что и витамин D. Возможно, именно поэтому у северных европейцев мутация, о которой идет речь, встречается чаще всего. Это пример генетической адаптации к взаимодействующим пищевым и климатическим факторам.
Еще несколько примеров. Эскимосы при традиционном питании обычно потребляют до 2 кг мяса в день. Переварить такие количества мяса можно лишь при сочетании определенных культурных (кулинарных) традиций, микрофлоры определенного типа и наследственных физиологических особенностей пищеварения.
У народов Европы встречается целиакия – непереносимость белка глутена, содержащегося в зернах ржи, пшеницы и других злаков. Она вызывает при потреблении в пищу злаков множественные нарушения развития и умственную отсталость. Заболевание в 10 раз чаще встречается в Ирландии, чем в странах континентальной Европы, вероятно, потому, что в ней пшеница и другие злаки традиционно не были основными продуктами питания.
У жителей Северноазиатского региона часто отсутствует фермент трегалаза, расщепляющий углеводы грибов. Эта наследственная особенность сочетается с культурной: в этих местах грибы считаются пищей оленей, не пригодной для человека.

Для жителей Восточной Азии характерна другая наследственная особенность обмена веществ. Известно, что многие монголоиды даже от небольших доз спиртного быстро пьянеют и могут получить сильную интоксикацию. Это связано с накоплением в крови ацетальдегида, образующегося при окислении алкоголя ферментами печени. Известно, что алкоголь окисляется в печени в два этапа: сначала превращается в токсичный ацетальдегид, а затем окисляется с образованием безвредных продуктов, которые выводятся из организма. Скорость работы ферментов первого и второго этапов (алкогольдегидрогеназы и ацетальдегидрогеназы) задается генетически. Для коренного населения Восточной Азии характерно сочетание «быстрых» ферментов первого этапа с «медленными» ферментами второго этапа. В этом случае при приеме спиртного этанол быстро перерабатывается в альдегид (первый этап), а его дальнейшее удаление (второй этап) происходит медленно. Такая особенность связана с сочетанием двух мутаций, влияющих на скорость работы упомянутых ферментов. Предполагается, что высокая частота этих мутаций (30–70%) есть результат адаптации к неизвестному пока фактору среды.
Приспособления к типу питания связаны с комплексами генетических изменений, не многие из которых пока детально изучены на уровне ДНК. Известно, что около 20–30% жителей Эфиопии и Саудовской Аравии способны быстро расщеплять некоторые пищевые вещества и лекарства, в частности амитриптилин, благодаря наличию двух или более копий гена, кодирующего один из видов цитохромов – ферментов, разлагающих чужеродные вещества, поступающие в организм с пищей. У других народов удвоение данного гена цитохрома встречается с частотой не более 3–5%, и распространены неактивные варианты гена (от 2–7% у жителей Европы и до 30% в Китае). Возможно, число копий гена увеличивается из-за особенностей диеты (использования больших количеств перца или съедобного растения тефф, составляющего до 60% пищевых продуктов в Эфиопии и нигде больше не распространенного в такой степени). Однако определить, где причина, а где следствие в настоящее время невозможно. Случайно ли увеличение в популяции носителей множественных генов позволило людям есть какие-то особые растения? Или, наоборот, употребление перца (или другой пищи, для усвоения которой необходим цитохром) послужило фактором отбора индивидов с удвоенным геном? Как тот, так и другой процесс могли иметь место в эволюции популяций.
Очевидно, что пищевые традиции народа и генетические факторы взаимодействуют. Употребление той или иной пищи становится возможным лишь при наличии определенных генетических предпосылок, а диета, ставшая традиционной, действует как фактор отбора, влияя на частоту аллелей и распространение в популяции наиболее адаптивных при таком питании генетических вариантов.
Традиции обычно меняются медленно. Например, переход от собирательства к земледелию и соответственно смена диеты и образа жизни осуществлялись на протяжении десятков поколений. Относительно медленно происходят и сопровождающие такие события изменения генофонда популяций. Частоты аллелей могут колебаться на 2–5% за поколение, из-за чего одни аллели постепенно накапливаются, а другие – исчезают. Однако другие факторы, например эпидемии, часто связанные с войнами и социальными кризисами, могут в несколько раз поменять частоты аллелей в популяции на протяжении жизни одного поколения за счет резкого снижения численности популяции. Так, завоевание Америки европейцами привело к гибели до 90% коренного населения, и эпидемии оказали большее значение, чем войны.
Устойчивость к инфекционным заболеваниям
Оседлый образ жизни, развитие земледелия и скотоводства, повышение плотности населения способствовали распространению инфекций и появлению эпидемий. Так, туберкулез – изначально болезнь крупного рогатого скота – человек приобрел после одомашнивания животных. С ростом городов заболевание стало эпидемически значимым, что сделало актуальной устойчивость к инфекции, также имеющей генетический компонент.
Наиболее подробно изученный пример подобной устойчивости –распространение в тропической и субтропической зонах болезни серповидноклеточной анемии, названной так из-за серповидной формы эритроцитов (определяется при микроскопическом анализе мазка крови). Эта наследственная болезнь обусловлена мутацией в гене гемоглобина, приводящей к нарушению его функций. Носители мутации оказались устойчивыми к малярии. В зонах распространения заболевания наиболее адаптивно гетерозиготное состояние: гомозиготы с мутантным гемоглобином погибают от анемии, гомозиготы по нормальному гену болеют малярией, а гетерозиготы, у которых анемия проявляется в мягкой форме, защищены от малярии.
Такие примеры показывают, что платой за повышенную адаптивность гетерозигот может быть гибель на порядок реже встречающихся гомозигот по болезнетворной мутации, которые неизбежно появляются при увеличении ее популяционной частоты.
Еще один пример генетической детерминации восприимчивости к инфекциям – так называемые прионные заболевания. К ним относится губчатая болезнь мозга рогатого скота (коровье бешенство), вспышка которого среди рогатого скота наблюдалась после появления новой технологии переработки костной муки, идущей на корм животным. Инфекция с очень небольшой частотой передается человеку через мясо больных животных. Немногие заболевшие люди оказались носителями редкой мутации, раньше считавшейся нейтральной.
Существуют мутации, защищающие от инфицирования вирусом иммунодефицита человека либо замедляющие развитие болезни после заражения. Две таких мутации встречаются во всех популяциях (с частотой от 0 до 70%), а еще одна – только в Европе (частота – 5–18%). Предполагается, что эти мутации распространились в прошлом в связи с тем, что обладают защитным эффектом 2 и в отношении других эпидемических заболеваний.
Развитие цивилизации и генетические изменения
Кажется удивительным тот факт, что питание бушменов – охотников-собирателей, живущих в Южной Африке, – оказалось полностью соответствующим рекомендациям ВОЗ по общему балансу белков, жиров, углеводов, витаминов, микроэлементов и калорий. Биологически человек и его непосредственные предки на протяжении сотен тысяч лет адаптировались к образу жизни охотников-собирателей.
Изменения традиционного питания и образа жизни отражаются на здоровье людей. Например, афроамериканцы чаще, чем евроамериканцы, болеют гипертонией. У северных народов, традиционная диета которых была богата жирами, переход на европейскую высокоуглеводную диету способствует развитию диабета и других заболеваний.
Преобладавшие ранее представления о том, что с развитием производящего хозяйства (земледелия и скотоводства) здоровье и питание людей неуклонно улучшается, сейчас опровергнуто. После появления земледелия и скотоводства значительное распространение получили многие заболевания, редко встречавшиеся у древних охотников-собирателей или вообще им неизвестные. Сократилась продолжительность жизни (от 30–40 лет до 20–30), в 2–3 раза увеличилась рождаемость и одновременно выросла абсолютная детская смертность, хотя относительный уровень ее, видимо не изменился: лишь 40% живорожденных детей доживали до репродуктивного возраста. Костные останки раннеземледельческих народов гораздо чаще имеют признаки перенесенной анемии, недоедания, различных инфекций, чем у доземледельческих народов. Лишь в Средневековье наступил перелом, и средняя продолжительность жизни стала увеличиваться. Заметное улучшение здоровья населения и снижение детской смертности в развитых странах связано с появлением современной медицины.
Сегодня для земледельческих народов характерны высокоуглеводная и высокохолестериновая диета, использование соли, снижение физической активности, оседлый образ жизни, высокая плотность населения, усложнение социальной структуры. Приспособление популяций к каждому из этих факторов сопровождается генетическими изменениями: адаптивных аллелей становится больше, а неадаптивных меньше, поскольку их носители менее жизнеспособны или менее плодовиты. Например, низкохолестериновая диета охотников-собирателей делает адаптивной для них способность к интенсивному поглощению холестерина из пищи, но при современном образе жизни она становится фактором риска атеросклероза и сердечно-сосудистых заболеваний. Эффективное усвоение соли, бывшее полезным при ее недоступности, в современных условиях превращается в фактор риска гипертонии. При рукотворном преобразовании среды обитания человека популяционные частоты аллелей меняются так же, как и при естественной адаптации.
Рекомендации врачей по поддержанию здоровья – физическая активность, прием витаминов и микроэлементов, ограничение соли и т.п. – по сути, искусственно воссоздают условия, в которых человек жил большую часть времени своего существования как биологического вида.
Вероятно, что определенные адаптации могли быть связаны и с коллективным образом жизни человека. Так, возросшая частота депрессий в современных обществах западного типа вызвана утратой поддержки родовой группы. В ряде исследований показано, что с разрушением родовой системы снижается выживаемость детей, повышается риск развития заболеваний. Согласно статистике, существенно различается частота депрессий в разных странах (в европейских она в пять раз выше), а частота шизофрении везде примерно одинакова. Как считают специалисты, генетическая детерминация депрессии довольно велика (30–40%). Можно предположить, что гены, ответственные за предрасположенность к депрессии, в обществах, где влияние коллектива еще велико, не столь опасны, как в обществе, где человек остается один на один со своими проблемами.
Итак, на формирование генофондов этнических групп влияет множество процессов: миграции и смешение народов, накопление мутаций в изолированных группах, адаптация популяций к условиям среды. Межпопуляционные (географические, языковые и иные) барьеры способствуют накоплению генетических различий, которые, однако, между соседями обычно не очень значительны. Географическое распределение этих различий отражает континуум меняющихся признаков и меняющихся генофондов. Генетические различия не подразумевают превосходства какой-либо расы, этнической или иной группы, образованной по какому-либо признаку (типу хозяйства или социальной организации). Напротив, они подчеркивают эволюционную ценность разнообразия, позволившую человечеству не только освоить все климатические зоны Земли, но и приспособиться к тем значительным изменениям среды, которые возникли в результате деятельности самого человека.
Литература
Генофонд и геногеография народонаселения России и сопредельных стран / Под ред. Ю.Г. Рычкова. – СПб., 2000.
Горбунова В.Н., Баранов В.С. Введение в молекулярную диагностику и генотерапию наследственных заболеваний. – СПб., 1997.
Лимборская С.А., Хуснутдинова Э.К., Балановская Е.В. Этногеномика и геногеография народов Восточной Европы. – М., 2002.
Степанов В.А. Этногеномика народов Северной Евразии – Томск, 2002.
Evolution in health and disease / Ed. S.C. Stearns. – N.Y., 1999.
Cavalli-Sforza L.L., Menozzi P., Piazza A. History and Geography of Human Genes, Princeton. – N.Y., 1994.
Кавалли-Сфорца Л.Л. Гены, народы, языки // В мире науки. 1992.
Уилсон А.К., Канн Р.Л. Недавнее африканское происхождение людей // В мире науки. 1992.
Боринская С.А., Хуснутдинова Э.К. Этногеномика: история с географией // Человек. 2002. № 1. С.19–30.
Хуснутдинова Э.К., Боринская С.А. Геномная медицина – медицина XXI века // Природа. 2002. № 12. С.3–8.
Геном человека: нити судьбы // Химия и жизнь. 1998. № 4. C.27–30.
Янковский Н.К., Боринская С.А. Наша история, записанная в ДНК // Природа. 2001. № 6. С.10–17.
Эти и другие научно-популярные статьи представлены на сайте www.vigg.ru в разделе «Программа «Геном человека».
Когда-то антропологи всего мира полагались только на заступ и лопату в своем стремлении достать из-под земли хронологические свидетельства развития высших приматов, появления Homo sapiens и колонизации этим видом всей планеты.
В XXI веке на смену археологическим находкам пришли данные всемирного генографического проекта, открывшего всему миру одновременно и существенное генетическое разнообразие всего человечества, и наметившего последовательность расселения людей по Земле. Например, выводы на основании раскопок древних поселений американских индейцев подтвердились и со стороны генетиков, и теперь всем доподлинно известно, что индейцы - потомки коренных сибиряков, переселившиеся на побережье северной Америки более 15 тысяч лет назад.
В то же время, попытки провести более детальный анализ последовательных волн миграции людей по планете на основании только лишь генетических данных пока даются с трудом. И зачастую антропологи стараются прибегать к генетическому анализу лишь для подтверждения спорных антропологических теорий, обосновать которые традиционными методами не удаётся.
Как результат, определённым налётом сомнения оказываются покрыты все выводы генетической антропологии - тем более что часто они оказываются весьма неожиданными. Однако огромное количество информации, скрытой в человеческих хромосомах, глупо было бы не использовать, и мы уже видим, как генетические методы находят всё большее использование в изучении истории развития человечества.
История миграции людей может обрасти массой новых подробностей уже очень скоро благодаря новому методу анализа генетического разнообразия,
разработанному специалистами из британского Оксфорда и американского Корнельского университета под руководством Дэниэла Фэлуша из Университета ирландского города Корк. Новая методика особенно удобна при сравнении коллективных геномов целых человеческих популяций и отдельных из них выборок, что позволяет сказать о генетическом родстве гораздо больше, чем прежде использованные приемы.
Новый статистический подход обещает установить временные рамки хотя бы основных миграционных развилок в расселении человека, а также размеры популяций, отделявшихся на этих развилках и воссоединявшихся позднее в современных народах. Свои результаты учёные опубликовали в PLoS Genetics.
У него три основных преимущества перед используемыми на сегодняшний день методиками.
Первое из них - приспособленность метода к учёту блочного копирования целых участков ДНК при смешивании человеческих популяций. Мутации при наследовании происходят двумя основными методами: точечным, когда изменению подвергаются отдельные «буквы» генетического кода; и блочным, когда копируются, переставляются или исчезают длинные участки ДНК. Используемые до сих пор методы учитывали лишь точечные мутации, не имея возможности надлежащим образом изучить сложную взаимозависимость между различными элементами генома. Такие методы сами генетики за глаза порой называют «плюшевой» генетикой.
Таким образом, по сути, в модель включаются не только существующие популяции, имеющиеся в реальных данных, но и синтетические популяции предков.
Конечно, на деле моделируются не реальные народы-предки, а их суррогаты, построенные из реальных данных на основании предполагаемой для каждого конкретного сценария иерархии популяций, отсортированных по отношению потомок — предок.
Как отмечают авторы, такая модель должна особенно хорошо работать в случае последовательного заселения людьми региона за регионом, так как «достраивать» при этом приходится меньшее число гипотетических популяций, и дело упирается только в выбор правильной последовательности «старшинства» народов. По современным представлениям, примерно так и происходило расселение людей: все мы вышли из Африки и затем колонизировали всё новые и новые территории.
Свою модель учёные протестировали на данных Human Genome Diversity Project (HGDP), опубликованных в 2006 году.
Они включают генетическую информацию о почти тысяче представителей 53 различных народов. Для каждого донора ДНК данные содержат информацию по 2 тысячам генетических маркеров; хотя метод Фэлуша и его коллег и приспособлен к обработке больших массивов данных по копированию отдельных участков генома, он может работать и с относительно «разреженными» данными, характерными для традиционного пока подхода.
На основании этих данных учёные выделили девять основных этапов колонизации нашей планеты. Генетики отмечают, что эти этапы не обязательно происходили в чётком хронологическом порядке, однако уверены, что правильно определили основных предков каждой популяции на каждом этапе.
Не удивительно, что заселение мира человеком происходит сначала по Африке - от сообществ охотников и собирателей на юге Африки в центр и на север континента и затем через Ближний Восток - в центральную часть Евразии. Проживающие здесь народы, в числе которых и российские адыгейцы, сильно перемешаны друг с другом, что, по мнению авторов, свидетельствует об отсутствии каких-то заметных «узких мест» в расселении людей в этот период.
Среди европейских народов «прародителями» модель выделяет французов, итальянцев и тосканцев, в генах которых нашёлся сильный сигнал от коренных жителей центральной Африки. В то же время, популяции из HGDP, проживающие на окраинах Европы, - сардинцы, баски, жители Оркнейских островов и русские - получили большое число генетического материала как от европейцев, так и с Ближнего Востока и из центра Евразии, абсорбируя пришельцев в Европу во время волн миграции, последующих за основной. У русских, под которыми подразумеваются жители севера европейской части России, особенно много предков из самых разных регионов Евразии.
При анализе выявились и весьма неожиданные детали.
Например, у якутов предки нашлись не только среди русских, что вряд ли кого-то может удивить, но и среди жителей Оркнейских островов, расположенных к северу от Шотландии.
Кроме того, жители Южной Америки позаимствовали некоторое количество генов от монголов, в то время как североамериканские индейцы произошли по большей части от народностей, населяющих ныне более северные районы Сибири, и не имеют монгольских корней. Впрочем, кровь сибиряков в жилах южноамериканцев остаётся доминирующей. Всё это может свидетельствовать всё-таки о нескольких независимых волнах миграции людей в Америку, вопреки большинству последних работ на этот счёт.
Ученые даже предлагают довольно правдоподобный сценарий развития этих событий. Популяции, которые первыми колонизировали северо-восточные регионы Азии, а впоследствии достигли и пересекли Берингов пролив, и чьи потомки в итоге заселили территории Южной Америки, были впоследствии вытеснены популяцией, более близкой к современному населению северо-восточного азиатского региона и в особенности монгольскому этносу.
Эта демонстрация силы нового статистического подхода наверняка потешит самолюбие разработчиков метода, так как она позволяет выявить древнее генетическое вливание в наследственную структуру популяции, даже если источник этого вливания на сегодняшний день не сохранился. В то же время, не следует забывать, что выводы Фэлуша и его коллег - не более чем наиболее правдоподобный сценарий. Впрочем, так работает почти вся современная наука, пытающаяся воссоздать прошлое, недоступное нашим приборам и органам чувств.
ГЕНЕТИКА ЧЕЛОВЕКА (демографические аспекты), раздел генетики, изучающий явления наследственности и изменчивости у человека. Материальной основой наследственности у человека, как и у других организмов, являются гены, расположенные в хромосомах и передающиеся в поколениях с помощью половых клеток. Каждый из генов представлен в организме дважды - один получен от отца, другой - от матери. В зависимости от различия или тождества унаследованных генов человек соответственно гетерозиготен (т.е. отцовский и материнский гены в данной паре не одинаковы) или гомозиготен (отцовский и материнский гены в данной паре одинаковы). Вероятность гомозиготности по совокупности генов из-за большого их числа (по разным оценкам, 105-106) крайне мала. Доля генов в гомозиготном состоянии у человека возрастает, если его родители имеют общих предков, от которых унаследовали идентичные гены. Такие случаи, регулируясь в человеческом обществе брачными традициями и законами, встречаются сравнительно редко, и, как правило, индивидуальный набор генов - генотип - формируется сочетанием родительских генов, происходящих из разных частей генофонда - общей совокупности генов популяции. Индивидуальное разнообразие набора генов огромно и образует биол. фундамент уникальности и неповторимости человеческой личности.
Один из важнейших разделов генетики человека - популяционная генетика человека. В отличие от популяций других видов популяция человека - объект действия и продукт не только естественно-исторического, но и общественно-исторического процесса. Воспроизводство генов человека, будучи, с одной стороны, сугубо биологическим процессом, с другой - социально обусловлено и неотделимо от демографического развития и воспроизводства народонаселения. Передача генетической информации в поколениях, ее распределение в пространстве расселения населения, изменение в ходе миграций, переселений, взаимодействий населения с окружающей средой - все эти движения генетического материала у человека связаны с демографическими процессами. Таким образом, популяционную генетику человека можно рассматривать как демографическую генетику, т. е. область взаимодействия генетики и демографии, исследующую генетические последствия демографических процессов.
Генофонд популяции, представленный в каждом поколении разнообразными генотипами, не остается постоянным во времени, т. к. благодаря дифференциальной рождаемости, смертности и миграции носители генов одного поколения в разной степени передают свои гены новым поколениям. Изменение популяционного генофонда, вызванное неодинаковым участием носителей разных генов в процессе воспроизводства, считается в общей теории популяционной генетики основным проявлением естественного отбора, который меняет структуру генофонда в сторону большего соответствия условиям среды. Другими факторами, действующими на изменения генофонда в популяциях человека, являются мутации, миграции и дрейф генов. Мерилом биологически нормальной, естественной скорости изменения генофонда является темп естественного мутационного процесса. Эффекту мутаций собственных генов генофонда эквивалентен эффект миграций генов из других популяций с существенно иным генофондом, т. к. при этом также возникают новые, ранее несвойственные популяции генотипы. Другое последствие регулярных миграций генов - стирание генетических различий между популяциями, потеря ими генетического своеобразия, возникшего в ходе самостоятельного развития и специфического приспособления к локальным условиям среды. Миграция генов осуществляется через миграцию их носителей. Роль миграции в истории развития народонаселения едва ли поддается однозначной оценке и трактовке, но некоторые ее генетические последствия очевидны, ибо значительная часть современного мирового населения представлена генетически смешанными популяциями. В несколько ином плане та же проблема возникает в связи с процессом урбанизации, вызывающим отлив населения из различных местных популяций и его прилив в центры урбанизации.
Даже в отсутствии мутаций, отбора, миграций генов (что почти невероятно) генофонд популяции все же сохраняет возможность изменяться. Происходит это в силу так называемого дрейфа генов, или генетико-автоматического процесса, - такого изменения генетической структуры популяции, которое вызывается случайными причинами, например, малыми размерами популяции. Дрейф генов наблюдается в численно небольших и преимущественно эндогамных популяциях - изолятах, где имеет место значительное несоответствие между потенциально всегда большим разнообразием возможных генотипов и малым числом реальных носителей генов. В силу малочисленности популяции в каждом поколении реализуется лишь малая часть возможных генотипов, и формирование генофонда нового поколения приобретает характер случайного выбора ограниченного числа генов из родительского генофонда. Популяционная генетика трактует дрейф генов как процесс, не зависящий от состояния среды. Вместе с тем именно на примере малых замкнутых популяций человека можно увидеть, что численность популяции определенным образом связана с уровнем общественно-экономического и культурного развития, а также с характером взаимодействия популяции со средой обитания. Таким образом, дрейф генов, зависящий от размера популяции, оказывается зависимым и от состояния общественной и природной среды.
Различные генетические процессы, рассмотренные выше порознь, в реальных популяциях представляют взаимосвязанные компоненты единого генетического процесса.
Основным источником информации о генетических процессах в населении является генетический полиморфизм, т. е. одновременное присутствие в популяции двух и более форм одного и того же наследственного признака или свойства. Он исследуется с помощью генетических маркеров - наследственных признаков, свидетельствующих о присутствии в генотипе человека тех или иных генов, обусловливающих эти признаки. Соответственно применяются разнообразные экспериментальные методы изучения генетических маркеров как источников информации о генотипах людей и генофондах популяций. Важную информацию о степени замкнутости и своеобразии генофонда в эндогамных популяциях, об уровне наследств, полиморфизма и т. п. позволяет получить генеалогия популяции, а также архивные и текущие записи актов гражданского состояния. Источником информации в генетике человека служат и такие сведения о населении, как его численность, брачность, семейная структура, рождаемость, смертность, расселение и пространств, структура, миграции. Гены, носителями которых являются современные поколения, дошли до них из глубокого прошлого, и поэтому генетика человека использует также данные археологии, этнографии и истории.
Генетические аспекты численности и демографической структуры населения . Население мира в целом, как и население, слагающее отдельные этносы, имеет сложную иерархическую популяционную структуру. В основании этой иерархии находятся элементарные популяции - простейшие единицы всей популяционной системы человечества. На нижнем уровне этой системы преобладают популяции сельского типа с численностью от десятков и сотен до тысяч человек. К этому же уровню относят и городские популяции с численностью от тысяч до миллионов человек. При различной численности и сельские, и городские популяции однотипны с том отношении, что лишены постоянных внутрипопуляционных барьеров, которые расчленяли бы их генофонд на относительно независимые и устойчиво воспроизводящиеся в поколениях части (в больших городах капиталистических стран в значительной степени сохраняется расчлененность генофонда в силу расовых, национальных, кастовых, религиозных и других различий). Число генов какого-либо типа в генофонде элементарной популяции вдвое больше числа составляющих ее людей. Однако с формированием генофонда следующего поколения связана лишь часть генов, носители которых - люди репродуктивного возраста. Из них не все вступают в брак, а из вступивших не все имеют детей или имеют разное их число и, наконец, не все дети доживают до репродуктивного возраста. Это означает, что даже гены, образующие ту часть генофонда, которая обеспечивает его воспроизводство, воспроизводят себя в разном числе копий. Чем меньшая часть генов родительского поколения воспроизвела себя в большем числе копий, тем больше генетические различия между поколениями популяции. В связи с этим генетически значимой является не общая численность популяции, а ее т. н. генетически эффективная численность - параметр, учитывающий все составляющие процесса воспроизводства - неравное соотношение полов, их неравную плодовитость, репродуктивную активность, ее продолжительность, различную в разных семьях выживаемость детей.
Отношение генетически эффективной численности к общей численности популяции зависит не только от биологических, но и от социальных факторов. В популяциях сельского типа это отношение составляет обычно около 1/3. В городских популяциях под выравнивающим влиянием социальной среды на репродуктивные показатели семей доля генетически эффективной численности может резко возрастать даже при сокращении воспроизводства и общего размера популяции. Размер популяции в свою очередь влияет на скорость генетических изменений в популяции: чем он численно больше, тем медленнее изменяется генетическая структура популяции. Поэтому там, где население состоит из большого числа элементарных популяций, наблюдаются значительные генетические различия между ними.
Генетические аспекты брачности . Многие моменты математического моделирования генетических процессов в популяциях связаны с принципом панмиксни (полной случайности образования брачных пар). В популяциях человека этот принцип реализуется с большими ограничениями. Общество, запрещая или поощряя, в зависимости от традиций и законов, родственные браки, регулирует степень панмиксии и воздействует на генетический процесс. В разных общественно-экономических и историко-культурных условиях различна и широта брачного круга, а следовательно, и уровень генетического разнообразия в нем. Ориентируясь, хотя бы частично, на психофизиологические (темперамент и т. д.), морфологические (тип телосложения, расовые особенности) и др. свойства, прямо или косвенно связанные с генотипом, человек тем самым производит неслучайный выбор из окружающего его разнообразия генотипов. Наибольшая избирательность наблюдается при близкородственных браках - инбридинге. Особенно высока его частота в изолятах, где преобладают внутренние (эндогамные) браки (их частота достигает почти 100%). В этом случае сама традиция эндогамии, несмотря на запрещение явно родственных браков, неминуемо порождает инбридинг. Чем меньше генетически эффективная численность изолята, тем с течением времени все более родственными становятся браки, и все более увеличивается генетическая однородность популяции. Уровень наследственного полиморфизма в таком изоляте сокращается, и популяция оказывается высокоадаптированной к узкому диапазону условий окружающей среды. Известны случаи, когда популяции, оказавшись на исторических окраинах мира и утратив в условиях изоляции некоторую долю наследственного полиморфизма (в частности, иммунологического), при контакте с пришлыми группами населения ценой больших потерь адаптировались к изменившейся эпидемиологической обстановке.
Широта брачного круга может сказываться и на таких признаках потомства, которые лишь частично определяются генотипом. С широтой брачного круга, т. е. с уровнем генетических различий родителей, до определенной степени связаны показатели физического развития детей, выносливости, устойчивости к стрессу, трудоспособности. В уровне этих различий, судя по влиянию на потомство, существует свой оптимум, означающий существование оптимума и в размерах круга брачных связей.
Генетические аспекты семейной структуры . Главный метод изучения закономерностей наследственной передачи признаков у человека - анализ распределения признаков у членов семьи в зависимости от степени их родства. Если признак, будучи генетическим маркером, не влияет на подбор супружеских пар, то доля родительских пар с определенным сочетанием маркирующих признаков обусловлена только частотой, с которой распространены в населении гены, кодирующие эти признаки. Например, группы крови человека, обозначаемые символами О(I), А(II), В(III) и AB(IV), кодируются тремя аллельными генами О, А и В. Распространение этих трех генов в мировом населении изучено особенно хорошо в силу их значимости для службы переливания крови. Семейная структура населения локальной ли популяции, народа, страны или мира в целом в отношении признака групп крови представлена 16 генетически различными типами супружеских пар. Частота каждого из этих типов всецело зависит от частоты трех аллельных генов А, В и О. Так, зная, что в Западной Европе эти гены представлены в генофонде в соотношении 26% (А), 6% (В), 68% (О), а в Южной и Восточной Азии в соотношении 20% (А), 20% (В) и 60% (О), можно заранее предсказать, что семья, где, например, мать группы крови О(I) и отец группы крови А (II), в Западной Европе составляют ок. 20%, а в Южной и Восточной Азии - около 10% всех супружеских пар. В семьях с супружескими парами этого типа часты случаи патологии повторных и многократных беременностей и родов на почве иммуногенетической несовместимости родителей. Социально значимые аспекты одного этого факта проявления генетических закономерностей в семейной структуре населения очевидны, Таким образом, существует связь между частотой, с которой гены представлены в генофонде населения, частотами генотипов людей и частотами генетически различных типов семей, передающих в следующее поколение определенную долю генов генофонда. Величина помех в передаче генетической информации в поколениях обратно пропорциональна числу детей в семьях и прямо пропорциональна степени различий семей по числу детей.
Родство в семье имеет определенную генетическую меру, определяющую долю общих генов у любых двух членов семьи, связанных общностью (даже отдаленной) происхождения. Наиболее распространенные типы родства могут быть выражены долей генов, унаследованных от общего предка. Это имеет значение в вопросах регулирования браков, в случае наследств, заболеваний и при медико-генетическом консультировании относительно риска заболевания, отмеченного в семье.
Генетические аспекты рождаемости . Индивидуальное развитие (онтогенез) человека находится под генетическим контролем, в наибольшей мере проявляющимся в ранние фазы - от образования зиготы (оплодотворенной яйцеклетки) до рождения и раннего детства. Такой контроль наиболее ясно выступает в явлении генетического определения (детерминации) пола системой двух так называемых половых хромосом (одной, полученной от отца, другой - от матери). Генетическая детерминация пола происходит в момент слияния родительских половых клеток и зависит от того, в каком сочетании половые хромосомы родителей оказались в новой зиготе. Генетически контролируется также взаимодействие плода с материнским организмом. По оценкам, не менее 10% всех зачатий оканчивается спонтанными абортами, обусловленными генетической несовместимостью матери и плода. Менее выраженная генетическая несовместимость сказывается в осложненном протекании беременности и родов. Наиболее известный пример проявления генетических факторов в беременности и рождаемости - резус-несовместимость матери и плода, а значит и супругов, возникающая в силу полиморфизма генов, контролирующих резус-группы крови. Этот вид генетической несовместимости особенно част в населении Европы, Неравная плодовитость различных генотипов способна в ряду поколений изменить генофонд путем преимущественного распространения одних и убыли других генов.
Генетические аспекты смертности . Одни гены, унаследованные человеком от родителей, функционируют на протяжении всей жизни, другие - лишь на определенном этапе онтогенеза, третьи, присутствуя в генотипе, могут так и не проявиться в фенотипе. Хотя все гены не меняются в течение жизни организма, в разных возрастных группах населения наблюдаются различия в частоте разных генотипов. Причина этого в неодинаковой выживаемости индивидуальных генотипов. Она наиболее очевидна, когда организм оказывается носителем так называемых летальных генов, приводящих к его гибели. В других случаях определенные генотипические комбинации в определенной среде в той или иной мере снижают жизнеспособность и тем самым влияют на индивидуальную продолжительность жизни. В популяциях, существующих в стабильной среде, повышенная смертность отдельных генотипов компенсируется их повышенной плодовитостью и, таким образом, не затрагивает генетических различий между поколениями. В иных условиях изменение частоты генотипов в популяции отражает направление ее генетической адаптации к изменениям окружающей среды. В человеческом обществе, прилагающем максимум усилий в борьбе со смертностью, генетические причины смертности в наибольшей мере сказываются на начальных этапах онтогенеза.
Причиной неодинаковой выживаемости генотипов является также различная степень устойчивости и подверженности людей заболеваниям, хотя преимущество одних генотипов перед другими в этом отношении не является ни абсолютным, ни постоянным. Неравная жизнеспособность разных генотипов - один из механизмов, поддерживающий наследственный полиморфизм в популяциях человека, причем величина различий в степени жизнеспособности обычно порядка одного - нескольких %. В некоторых случаях (при появлении в среде патогенного фактора) соотношение в выживаемости генотипов достигает десятков %. Наиболее известный пример такого рода связан с серповидно-клеточной анемией - болезнью, первопричина которой в мутации одного из генов, кодирующих синтез гемоглобина. Если у какого-либо индивида в обеих гомологичных хромосомах присутствует мутантный ген (HbS), то такой индивид страдает тяжелой анемией и, как правило, не доживает до зрелости. Таким образом, при генотипе HbS HbS весь гемоглобин принадлежит к аномальному типу и разница в выживании такого генотипа по сравнению с нормальным НbA НbA составляет практически 100%. Однако в условиях тропической Африки и субтропического Средиземноморья разница в выживании меньше 100% в силу низкой устойчивости нормального генотипа НbA НbA к поражению малярийным плазмодием, для развития которого аномальный гемоглобин представляет менее подходящую среду, чем нормальный. Наиболее жизнестойки индивиды с генотипом HbA HbS, у которых ген НbA обеспечивает образование нормального гемоглобина, а ген HbS защищает от поражения малярийным плазмодием.
Генетические аспекты воспроизводства населения . В понятиях генетики человека воспроизводство населения есть воспроизводство генов человека в ходе смены поколений. Генетически ключевыми единицами в воспроизводстве населения являются элементарные популяции, дифференцированный рост которых в ходе воспроизводства ведет к неодинаковому распространению в населении генов из того или иного генофонда. Поскольку элементарные популяции человека не существуют вне этносов, в их неравном воспроизводстве отражено неравное же воспроизводство этнических генофондов, необратимо меняющее генетические свойства населения, что сказывается не только в постепенном изменении физического облика поколений, но и в нарушении устойчивости к патогенным факторам среды. Генетически значимая единица времени в воспроизводстве - поколение. В воспроизводстве генов нового поколения участвуют обычно 2 из 3-4 одновременно сосуществующих поколений, что сокращает возможность резких изменений в генетической структуре нового поколения и обеспечивает большую генетическую преемственность между поколениями. Охрана генетических механизмов воспроизводства - ключевое условие поддержания нормального физического состояния поколений. Посредством воспроизводства населения из отдаленного прошлого в настоящее и будущее передаются древние гены, обусловливающие физическое и психическое единство и целостность человечества во всем его многообразии. Воспроизводством могут быть подхвачены и новые гены, возникающие в результате мутаций. Систематический контроль за частотой генных мутаций - один им методов оценки генетического состояния среды и нормального хода воспроизводства.
Генетические аспекты миграции и расселения населения . Миграция населения приводит к миграции генов человека. Миграция генов в популяцию, изменяя генофонд, формируя новые генотипы, меняя установившиеся в поколениях соотношения приспособлеyнностей генотипов, усиливая дифференциальную плодовитость и выживаемость, выступает как фактор, воздействующий на течение генетического процесса в популяции. Различают интенсивность и генетическая эффективность миграции. При одинаковой интенсивности генетическая эффективность миграции тем больше, чем больше генетическое своеобразие популяций, обменивающихся генами, а генетическое своеобразие тем больше, чем больше размерностей у пространства, в котором происходит миграция. Социальная природа человека способствует увеличению числа размерностей миграционного пространства свыше двух - трех, свойственных популяциям других организмов, однако она же создает условия и стимулы к преодолению этого пространства, разделяющего популяции. Негритянское гетто Нью-Йорка, азиатские кварталы Сан-Франциско, Ист-Энд и Уэст-Энд Лондона, Замоскворечье и Белый город дореволюционной Москвы - все это не столько территориально, сколько социально разобщенные пространства, в которых происходят миграции генов, часто однонаправленные (например, от белых американцев к черным, но почти никогда - обратно). Преодоление такого пространства оказывается часто более трудным, чем преодоление географических расстояний. Когда миграция перестает зависеть от любого рода расстояний между популяциями, ее влияние, нивелирующее генетическое разнообразие популяций, становится максимальным. В популяциях, генетическое развитие в которых протекает по стационарному типу, миграция выступает в качестве фактора, регулирующего уровень генетического разнообразия, необходимый для поддержания адаптационной пластичности населения в изменяющейся окружающей среде. Этот уровень оказывается единым для коренного населения разных континентов и указывает на то, что в ходе истории был выработан оптимальный режим для всех генетических процессов в населении. Такой режим обеспечивает распределение всего эволюционно накопленного генетического разнообразия населения на внутрипопуляционные и межпопуляционные компоненты примерно в соотношении 90% а 10%. Такое же соотношение обнаружено в различных популяциях животных и растений, что подчеркивает его уникальную эволюционную важность для выживания. Соотношение внутри- и межпопуляционного генетического разнообразия легко вычисляется из демографических данных о миграции и численности населения. Поэтому эти данные могут служить для генетической оптимизации миграции населения и демографических процессов в целом.
В череде поколений относительно изолированного автохтонного развития генофонд каждой популяции и каждой группы мирового населения приобретает отличительные черты. Так сложились, например, существенно разные генофонды населения на территории СССР к 3ападу и Востоку от Урала, проявляющиеся даже в антропологических типах. Вместе с тем генофонд коренного населения обширного района между Волгой и Обью являет промежуточные черты, сложившиеся в результате длившегося тысячелетиями просачивания и миграций генов между европейскими и азиатскими частями общего генофонда древнего населения нашей страны. В эпоху Великого переселения народов миграция масс населения центрально-азиатского и южно-сибирского происхождения привела к широкому распространению генов из азиатского генофонда среди населения Европейской части СССР и Европы в целом. Последствия этих миграционных процессов древности до сих пор отражены в геногеографии населения Северной Евразии. Считается, что вызванная этими миграциями перестройка генофонда населения Европы сопровождалась изменением адаптационных свойств генотипов людей. Это проявилось, в частности, в распространении в населении Европы резус-несовместимости матери и плода, которая не встречается в Азии и очень редка на крайнем 3ападе Европы у басков. Одно лишь это «эхо» древних демографических процессов, нарушивших естественный ход и направление генетического развития населения Европы, требует сегодня особых профилактических мероприятий по охране материнства и детства. В геногеографии мирового населения отражены и многие другие события мировой демографической истории.
Обращенная в будущее, генетика человека дает ключ к пониманию и оценке возможных отдаленных генетических последствий современных демографических процессов.
Ю.Г. Рычков.
Демографический энциклопедический словарь. - М.: Советская энциклопедия. Главный редактор Д.И. Валентей. 1985.
Литература:
Ниль Дж., Шэлл У., Наследственность человека, пер. с англ. М. 1958; Штерн К, Основы генетики человека, пер. с англ., М. 1965; Маккьюсик В., Генетика человека, пер. с англ., М. 1967; Бочков Н. П, Генетика человека, М. 1978; Л и Ч., Введение в популяц. генетику, пер. с англ., М. 1978; Беляев Д. К., Совр. наука и проблемы исследования человека, «Вопросы философии», 1981, № 3.
Sforza L. L., Воrimer W. F., The genetics of human populations, S. F., 1977.
Источник: http://vivovoco.rsl.ru/VV/JOURNAL/VRAN/03_07/ETHNOGENE.HTM
Э.К. Хуснутдинова
ЭТНОГЕНОМИКА И ГЕНЕТИЧЕСКАЯ
ИСТОРИЯ НАРОДОВ ВОСТОЧНОЙ ЕВРОПЫ
Э. К. Хуснутдинова
Хуснутдинова Эльза Камилевна
- член-корреспондент Академии наук Республики Башкортостан,
заведующая отделом Института биохимии и генетики Уфимского научного центра РАН.
Эволюция человеческих сообществ (популяций), их происхождение, родство, историческое развитие всегда были в центре внимания многих наук. Для решения этих проблем необходимо исследовать множество признаков в большом числе популяций и этнотерриториальных групп. В качестве таких признаков можно использовать вариабельность структуры биополимеров (белки. нуклеиновые кислоты). Раньше основное внимание уделяли полиморфным белкам. Подлинный переворот в исследованиях произошел при появлении нового типа маркеров, основанных на геномных ДНК.
Геном человека, состоящий примерно из 3 млрд. нуклеотидных пар, расшифрован почти полностью. Однако само по себе близкое завершение этой гигантского по замыслу и грандиозного по реализации международного научного проекта отнюдь не означает, что процесс познания генома завершен. Уже сейчас очевидно, что не существует какого-то "усредненного" генома человека: каждый геном, как и каждый человек, сугубо индивидуален. Эта индивидуальность проявляется на уровне не только отдельной личности, но и этнических групп, отдельных сообществ и рас.
Различия между двумя людьми на уровне ДНК составляют в среднем один нуклеотид на тысячу. Именно этим обусловливаются наследственные индивидуальные особенности каждого человека. Заметим, что различия между ДНК человека и шимпанзе - его ближайшего сородича в животном мире - на порядок больше.
ОСНОВНОЙ ИНСТРУМЕНТ ЭТНОГЕНОМИКИ
Для исследований геномов людей используют разные типы ДНК-маркеров: расположенные на парных хромосомах (аутосомные), на митохондриальной ДНК и на непарной Y-хромосоме. Маркеры на парных хромосомах наследуются по обеим - женской и мужской - линиям, в них представлена подавляющая часть генома человека. Таким образом, у нас в руках оказались маркеры, позволяющие изучать сочетанную изменчивость, одновременно привносимую и с отцовской, и с материнской стороны. Все маркеры ДНК характеризуют сообщества в целом, не выделяя генетического вклада каждого из полов. Используя определенные типы полиморфизма ДНК, можно оценить те или иные временные события, происходившие в истории данной популяции.
Особую роль играют маркеры митохондриальной ДНК (мтДНК) и ДНК Y-хромосомы, поскольку они помогают проследить генетическую историю человечества отдельно по женской и мужской линиям. Митохондриальная ДНК передается потомкам только от матери, так как митохондрии находятся в цитоплазме клетки, а цитоплазма потомка (зиготы) образуется за счет цитоплазмы материнской яйцеклетки. Если два человека имеют общего предка женского пола, то по различиям их мтДНК можно судить о том, сколько поколений отделяет их от жившей столетия или тысячелетия назад общей пра-... прабабушки. Аналогично изучение ДНК Y-хромосомы позволяет проследить эволюционные траектории по отцовской линии, поскольку Y-хромосома передается только от отца к сыну. Оба типа полиморфизма ДНК дополняют друг друга, давая раздельную информацию об отцовском и материнском вкладе в этническую историю и эволюцию популяций.
Передаваясь из поколения в поколение только по одной из родительских линий и не участвуя в рекомбинации (обмен участками гомологичных хромосом в процессе мейоза), оба типа полиморфизма позволяют, по крайней мере теоретически, реконструировать генетические события от наиболее популярных предков современного человека - "Y-хромосомного Адама" и "митохондриальной Евы" - до современных популяций. Полиморфизм маркеров мтДНК и Y-хромосомы определяется факторами микроэволюции (миграция, отбор, мутации). Однако характер их вариабельности по-разному отражает действие и результат этих процессов.
Уровень разнообразия геномов представителей какого-либо биологического вида зависит, во-первых, от разнообразия геномов прародителей вида и, во-вторых, от скорости накопления случайных "ошибок" (мутаций), возникающих при "переписывании" клеткой генетических текстов, и еще от того, как долго существует данный вид. При сравнении генетических текстов Y-хромосомы (или мтДНК) разных людей по присутствию в них одинаковых мутаций можно выявить общего предка. Согласно современным представлениям, скорость накопления мутаций в ДНК относительно постоянна, большинство мутаций нейтральны, так как не затрагивают значимые, смысловые участки генома. Поэтому они не отсеиваются отбором и, раз появившись, передаются из поколения в поколение. Сравнивая два родственных генетических текста, по количеству различий между ними можно установить время появления как мутаций, так и общего предка по мужской или женской линиям.
Изучение полиморфизма ДНК позволяет выявлять значительные внутри- и межпопуляционные различия в частотах полиморфных маркеров ДНК во многих географических районах мира, что стало одной из важнейших характеристик генетической структуры человеческих сообществ. За последнее десятилетие генетиками собраны и проанализированы коллекции мтДНК и Y-xpoмосом представителей народов почти всего мира. По ним восстановлена последовательность и время появления мутаций в ДНК человека.
ДНК-маркеры - эффективный инструмент для исследования гаплотипов - сочетаний аллелей тесно сцепленных полиморфных локусов. Аллель - одна из возможных альтернативных форм гена, а локус - область локализации гена в хромосоме или молекуле ДНК. Такие хромосомные участки (гаплотипы) весьма невелики по размерам, поэтому очень редко рекомбинируют. Они ведут себя как единые блоки, мало меняющиеся во времени и поэтому имеющие довольно древнее происхождение. Таким образом, размер сохранившегося неизменным гаплотипа может служить мерой времени, которое прошло от какого-то момента в прошлом. В общем случае суть анализа гаплотипов состоит в поиске сцепления между собой локусов вследствие эффекта основателя. Анализ частоты и возраста появления в популяции гаплотипа позволяет проследить его историю, а вместе с ней и события, сопутствующие его распространению. Очевидно, что такие данные представляют большой интерес для изучения истории современных народов, характеристики генофондов и оценки основных направлений эволюции всего человечества.
В 80-90-х годах прошлого века шло интенсивное накопление знаний об изменчивости мтДНК человека, были описаны основные расовые и по-пуляционно-специфические типы мтДНК. Глобальный скрининг всех основных расовых групп человечества по полиморфизму митохондриаль-ного генома позволил выявить наиболее древние мутации - ключевые для определения расоспеци-фических кластеров. Установлено, что митохонд-риальные геномы представлены комбинациями расовых групп типов мтДНК, каждая из которых ведет происхождение от единственного основателя.
Сейчас изменчивость митохондриального генома изучается на качественно новом уровне. Анализ однонуклеотидных замен мтДНК, рыяв-ляемых классическим методом анализа полиморфизма ДНК, в сочетании с изучением нуклеотид-ной последовательности гиперизменчивой области мтДНК позволяет проводить комплексную оценку полиморфизма мтДНК в популяциях человека. Такой подход выявил важную для изучения молекулярной эволюции особенность мито-хондриальной ДНК: определенным расовым группам типов мтДНК, ключевые мутации которых расположены в различных участках молекулы, соответствуют вполне определеные типы нуклеотидных последовательностей гипервариа-бельного участка мтДНК. Поскольку существует неравновесие по сцеплению между мутациями в митохондриальном геноме, молекулу ДНК можно рассматривать как один локус, представленный множеством аллелей, определенные группы которых соответствуют определенным группам сцепления между конкретными мутациями. Эти два подхода положены в основу классификации и эволюции линий всего митохондриального генома современного человечества.
ГЕНОМНЫЙ КЛЮЧ К ПРОБЛЕМЕ ПРОИСХОЖДЕНИЯ НАРОДОВ
Первым приложением ДНК-маркеров к проблеме происхождения и расселения человечества стали выполненные Канном с коллегами исследования митохондриальной ДНК представителей различных рас - африканцев, европейцев, азиатов, австралийцев и жителей Новой Гвинеи. По количеству замен нуклеотидов в мтДНК была определена степень родства различных групп людей и построено эволюционное древо человечества (рис. 1). Самая ранняя точка ветвления на древе отделяет от остальных людей группу африканцев, что указывает на африканское происхождение Homo sapiens . Именно в Южной Африке найдены самые древние мутации у бушменов и готтентотов и зарегистрировано самое высокое разнообразие мтДНК.
Рис. 1.
Эволюционное древо человечества.
Числа соответствуют количеству исследованных популяций;
проценты - уровню внутрипопуляционного разнообразия,
оно самое высокое (0.6%) в африканских популяциях и самое низкое (0.2%) - в европейских
Митохондриальные ДНК у населения других континентов менее разнообразны, и сравнение их с мтДНК аборигенов Южной Африки показало, что они возникли как мутационные изменения африканских типов после того, как человечество распространилось за пределы этого континента. Второй вывод касался времени коалесценции (расхождения) митохондриальной ДНК. По дате отделения ветви шимпанзе (5-7 млн. лет назад), приняв темп мутационной дивергенции равным 2-4% за 1 млн. лет, Канн с коллегами вычислили время существования последней предковой мтДНК, общей для всех ныне живущих людей, - примерно 185 тыс. лет назад.
Последующие работы подтвердили африканские корни древа мтДНК современного человечества, хотя и остаются еще отдельные спорные моменты. По незавивимым оценкам нескольких групп исследователей, "митохондриальная Ева" жила в период резкого сокращения численности наших предков (до 10 тыс.), вызванного, по-видимому, изменениями климата, - 185 тыс. лет назад. Именно этот период считают временем появления Homo sapiens как биологического вида. Сравнительное исследование митохондриальной ДНК разных популяций современных людей позволило выдвинуть предположение, что еще до выхода из Африки (около 60-70 тыс. лет назад) предковая популяция разделилась по крайней мере на три группы, давшие начало трем расам - африканской, монголоидной и европеоидной .
Вскоре были построены генеалогические древа по данным изучения ДНК Y-хромосомы . Исследование небольшого участка Y-хромосомы свидетельствует о возможно гораздо более позднем происхождении "Y-хромосомного Адама" - 140-175 тыс. лет назад. Результаты всех исследований указывают на его африканское происхождение. Различия между оценками, базирующимися на мтДНК и Y-хромосоме, могут быть объяснены как несходством демографической истории популяций по мужской и женской линиям, различным поведением женщин и мужчин при переселениях, завоеваниях и колонизациях, так и различиями самих геномов, например, в интенсивности отбора вариантов мтДНК и Y-хромосомы.
Гипотезу африканского происхождения современного человека подтверждает и наибольший уровень наследственного разнообразия в Африке по сравнению с другими континентами, а также малые различия между популяциями (на долю межпопуляционного разнообразия приходится 10-15% геномной вариабельности), что отражает недавнее происхождение биологического вида . В целом массив геномных данных наиболее соответствует гипотезе недавнего африканского происхождения современного человека и доказывает справедливость монофилетической гипотезы. В то же время генетические данные не являются исчерпывающим и бесспорным доказательством этой гипотезы.
На основе распределения у разных народов частот различных мутаций в Y-хромосоме и мтДНК составлена карта расселения людей с африканской прародины . Первые волны расселения человека современного типа прошли из Африки через Азию в Австралию и Европу. Удивительно, что время расселения человека по разным континентам соответствует датировке сделанных на этих континентах археологических находок. Например, появление человека в Австралии и Новой Гвинее датируется 50-60 тыс. лет назад, согласно генетическим данным. То же самое время показывает анализ изотопного состава химических элементов в археологических находках. В Центральной и Юго-Восточной Азии люди появились примерно 70 тыс. лет назад. Заселение Европы произошло позже, около 35-40 тыс. лет назад. Наиболее спорны оценки времени заселения Америки. Люди появились там гораздо позже, чем на других континентах, потому что нужно было пересечь Сибирь, добраться до Чукотки и воспользоваться тем моментом, когда уровень моря в период оледенения позволял перейти нынешний Берингов пролив. Случилось это в промежуток времени от 15 до 35 тыс. лет назад. Позже, под натиском ледника, палеолитические европейцы несколько раз отступали на юг и юго-восток, возможно, даже возвращались обратно в Африку, о чем свидетельствуют результаты исследования гаплотипов Y-хромосомы в популяциях Африки . Сравнивая спектр мутаций в ДНК современных европейцев и их азиатских соседей, удалось установить, что 10-20% генов было привнесено в Европу неолитическими переселенцами с Ближнего Востока около 10 тыс. лет назад. Вместе с ними в Европе появилось земледелие.
Разные расы и народы возникли после разделения предковых популяций. Эволюция вновь образовавшихся популяционных групп шла независимо. В каждой из них накапливались свои мутации, увеличивалась генетическая дистанция между группами. Сообщества приспосабливались к климатическим и географическим условиям, типу питания. В изолированных группах независимо протекала эволюция языка и культуры.
На формирование современных народов влияли не только процессы разделения популяций, поскольку народы могут образовываться при смешении нескольких исходных сообществ с разной расовой и языковой принадлежностью. Тогда возникает генетически разнородная этническая общность, но с единым типом культуры и общим языком. В связи с этим все большую актуальность приобретает изучение генетической истории популяций отдельных регионов, расово-этнических групп, генетической родословной современных этносов.
ЭТНОГЕНОМИКА НАРОДОВ ВОЛГО-УРАЛЬСКОГО РЕГИОНА
В Волго-Уральском регионе столкнулись две волны расселения: европеоидная и монголоидная. Находясь на границе двух частей света - Европы и Азии, этот регион на протяжении исторически длительного времени был местом взаимодействия многих этнических слоев. В формировании народов края известна роль угров Западной Сибири, финнов севера Восточной Европы, индо-иранцев Ближнего Востока, тюрков Южной Сибири и Алтая, а позднее кочевых татаро-монголь-ских племен и славянских народов Центральной и Западной Европы. Вплоть до XVI в. Среднее Поволжье было подлинным "котлом", в котором перемешались многие этнические группы. Следы взаимопроникновения хранят гены проживающих здесь народов.
Современные популяции Волго-Урала крайне неоднородны по этнолингвистической структуре. Сейчас здесь проживают представители финно-угорской ветви Уральской языковой семьи (удмурты, марийцы, мордва, коми), тюркской ветви Алтайской языковой семьи (башкиры, татары, чуваши) и восточно-славянской ветви Индоевропейской языковой семьи (русские). С целью познания факторов формирования генофонда, генетической реконструкции родства и происхождения современных народов Волго-Урала изучены 17 полиморфных аутосомных ДНК-локусов, определена нуклеотидная последовательность ДНК гипервариабельного участка митохондриального генома, выполнен анализ полиморфизма 24 локусов Y-хромосомы. В ходе исследований проанализированы геномы 1500 индивидов из 8 популяций (башкиры, татары, русские, чуваши, удмурты, коми, мордва, марийцы).
Полиморфизм аутосомных ДНК-локусов. Анализ полиморфизма 17 аутосомных ДНК-локусов в популяциях Волго-Уральского региона свидетельствует об их неоднородности и высокой информативности для популяционно-генетических исследований. Оценка степени генного разнообразия (Gst) народов этого региона показала, что их генофонд не только географически занимает промежуточное место между европейскими и сибирскими генофондами. Уровень генетических различий между этносами Волго-Урала (Gst = 1.91%) занимает как бы промежуточное положение между европейскими (Gst = 1.18%) и сибирскими народами (Gst = 5.84%), тяготея к европейскому типу (рис. 2). Изучение соотношения европеоидного и монголоидного вкладов в генофонд народов Волго-Уральского региона по данным полиморфизма ДНК-локусов выявило большую долю европеоидности. Самая высокая доля европеоидности обнаружена в популяциях мордвы (89.4%) и коми (81.0%), самая низкая - в популяциях башкир (62%) и марийцев (53%) . Это согласуется с данными антропологии и археологии.
Рис. 2 . Уровень генетических различий между этносами Сибири, Волго-Уральского региона и Европы
Полиморфизм мтДНК. Чтобы судить о генетическом разнообразии народов Волго-Уральского региона по материнской линии, была определена последовательность гипервариабельного участка митохондриальной ДНК (377 нуклеотидов) и проведен анализ полиморфизма мтДНК с использованием 26 эндонуклеаз (ферментов рестрикции). Полученные данные анализировались методом так называемых медианных сетей. Медианная сеть - один из графических вариантов представления данных о полиморфизме ДНК, который позволяет проследить дивергенцию митохондриального генома от предковой ДНК. В качестве предкового выбирается гаплотип, который широко распространен в данной популяции и включает наиболее частые аллели, характерные для данного этноса. В процессе такого анализа в единую медианную сеть соединяются посредством линий сотни и тысячи соседствующих, наиболее близких индивидуумов. В этой сети узловые соединения совпадают со специфическими гаплотипами, определенными по характерным мутациям изучаемого генома.
При сравнительном изучении мтДНК у народов Волго-Уральского региона выявленные гап-лотипы были отнесены к определенным линиям в соответствии с классификацией Макуолли . Для большинства расовых и географических групп установлена специфичность типов мтДНК (рис. 3). Более 90% всех типов мтДНК народов Западной Евразии принадлежит к линиям Н, J, V, 1, К, Т, U, W и X, которые рассматриваются как потомки европейского генного пула, существовавшего в верхнем палеолите. Митохондриальный геном азиатских популяций включает два суперкластера М и N, которые соединяются в африканском макрокластере L3. Последний рассматривается в качестве общего предка для всех неафриканских популяций мира. Около 50% линий мтДНК коренных жителей Азии принадлежит к суперкластеру М, который подразделяется на линии С, Z, D, G, Е и др. . Суперкластер N в Азии делится на несколько линий: А, Y, В, F.
Рис. 3. Медианная сеть, демонстрирующая полиморфизм мтДНК в популяциях Европы, Азии и Африки
Линии D, С, Z, G суперкластера М, линии A, Y суперкластера N и линии В, F суперкластера R характерны для азиатских популяций; линии 1, X, W суперкластера N и линии V, Н. Т, J, U, К суперкластера R - для европейских; линии LI, L2, L3, М1 и U6 принадлежат африканским народам
Большинство типов мтДНК народов Волго-Уральского региона соответствует линиям мтДНК Европы и Ближнего Востока, что свидетельствует об общих предковых линиях мтДНК, специфичных для европейцев . В целом среди изученных нами популяций частота европейских типов мтДНК оказалась наиболее высокой у мордвы, коми-зырян и русских. С другой стороны, уровень распространения линий мтДНК, специфичных для Восточной Евразии, также достигает больших значений, что ранее не было показано для Западной Европы. Высокая частота линий G, D, С, Z и F в некоторых этнических группах, как тюркских (башкиры), так и финноугорских (удмурты, коми-пермяки), указывает на значительное участие сибирского и центральноазиатского компонента в этногенезе народов Волго-Уральского региона.
Самостоятельный интерес вызывает высокая частота азиатской линии F (6%) у башкир. Эта линия характерна для народов Средней Азии - казахов, уйгуров и монголов, и мы можем предположить, что, во-первых, существенную роль в формировании данной этнической группы сыграл среднеазиатский компонент и, во-вторых , популяция башкир длительное время находится в изоляции от своих ближайших соседей. У других тюркоязычных и финноугорских популяций Волго-Уральского региона частота азиатских линий низкая. Поскольку очевидных географических барьеров как между тюркскими этносами, так и между финноугорскими популяциями не существует, можно сделать вывод о различной демографической истории башкир по материнской линии внутри упомянутых языковых семей. Выборки башкир и удмуртов по совокупности всех полученных по материнской линии данных можно охарактеризовать как этносы, имеющие в прошлом период резкого роста численности в условиях относительной изоляции. Анализ митохондриального генома татар, чувашей, марийцев, мордвы, коми и русских, скорее всего, отражает процессы продолжающейся интенсивной метисации в условиях сохранения постоянного популяционного размера. В целом, медианные сети показывают смешение и взаимопроникновение гаплотипов мтДНК, что свидетельствует как о тесных этногенетических контактах изученных этносов, так и о единой генетической основе населения Волго-Уральского региона по материнской линии.
В распространении типов мтДНК у народов Волго-Уральского региона ведущую роль играют факторы этнокультурной и территориальной близости или отдаленности, но не лингвистические барьеры. Это означает, что по материнской линии финноугорские народы имеют большее сходство с их непосредственными тюркскими соседями, чем с родственными в языковом отношении балтофинскими народами.
Анализ европеоидного и монголоидного вклада в материнские генетические линии народов Волго-Уральского региона не выявил корреляции языка и геномного состава этнических групп. На языках тюркской группы, привнесенных из Азии, говорят не только башкиры (65% монголоидности), но и татары и чуваши, у которых преобладает европеоидный генетический компонент. В остальных популяциях региона вклад монголоидного компонента составляет от 12% у русских до 20% у удмуртов. Русские, проживающие на территории этого региона, имеют 10-12% монголоидных типов мтДНК, а русские из Рязанской и Курской областей - только 2-3%. Это можно объяснить смешением русских с тюркоязычными народами на территории Волго-Уральского региона.
Интересно, что некоторые материнские линии у разных народов, например, у русских, татар и марийцев, оказались общими. Это показывает глубокое родство народов, говорящих на разных языках, придерживающихся разных религий и традиций.
Сравнительный анализ типов мтДНК в 18 популяциях Евразии, включая популяции Волго-Уральского региона (гагаузов, турков, татар, башкир, чувашей, карачаевцев, кумыков, азербайджанцев, узбеков, казахов, киргизов, ногайцев, уйгуров, шорцев, тувинцев, долган, якутов), которые относятся к тюркской ветви Алтайской языковой семьи, позволил установить западно-восточный градиент увеличения частоты азиатских линий мтДНК на расстоянии 8000 км: от 1% у гагаузов из Молдавии до 95% у якутов и 99% у долган (рис. 4). Кроме того, установлено, что лингвистическое сходство популяций играет меньшую роль, чем географическая близость или отдаленность популяций.
Рис. 4.
Результаты сравнительного анализа типов мтДНК в 18 популяциях Евразии
Отчетливо виден западно-восточный градиент увеличения частоты азиатских линий мтДНК
Один из наиболее важных аспектов анализа митохондриального генома - оценка времени коалесценции (расхождения, дивергенции) линий мтДНК в пределах каждой линии. Безусловно, на временные оценки будут влиять различные факторы формирования разнообразия мтДНК: объем выборки, миграция населения, резкий рост численности, феномен "бутылочного горлышка" - сильное сокращение численности наших предков, вызванное, по-видимому, изменением климата, и т.д. Тем не менее оценка времени дивергенции линии возможна при обнаружении предковых гаплотипов.
По ориентировочным оценкам, возраст дивергенции линий, выявленных у народов Волго-Уральского региона, варьировал от 273 ± 57 тыс. лет для азиатской линии Z до 22.76 ± 5.250 тыс. лет для линии С. Возраст дивергенции самой крупной европейской линии Н определен в 20.036 ± 4.250 тыс. лет, что соответствует археологическому времени повторной экспансии населения на территории Урала в постледниковый период. Используя данные о числе мутационных замен и скорости накопления мутации для гипервариабельного участка мтДНК, равной одной мутационной замене за 20.18 тыс. лет, мы получили среднее значение времени дивергенеции мтДНК для народов Волго-Уральского региона. Оно составляет 49.60 тыс. лет назад, что соответствует периоду расселения человека на европейском континенте в эпоху верхнего палеолита.
Полиморфизм ДНК Y-хромосомы. Анализ Y-хромосомы вошел в арсенал методов эволюционной генетики лишь в самое последнее время, когда был найден ряд высокоинформативных полиморфных локусов в ее нерекомбинантной части. Генетические свойства Y-хромосомы, такие как передача только по отцовской линии, отсутствие рекомбинации, малая эффективная численность пула Y-хромосом по сравнению с аутосомами (в четыре раза меньше, чем у аутосом), позволяют прослеживать по гаплотипам Y-хромосомы отцовские линии, представляющие собой последовательную "запись" мутаций в ряду поколений. По сравнению с митохондриальным геномом, насчитывающим 16.5 тыс. пар нуклеотидов. Y-хромосома, размер которой оценивается приблизительно в 60 млн. пар нуклеотидов, дает в руки исследователей потенциально более мощное "оружие".
Если предшествующие работы, посвященные анализу Y-хромосомы в популяциях России, основывались преимущественно на анализе 9 маркеров, то для изучения и сравнения генетического разнообразия отцовских линий в популяциях Волго-Уральского региона использовались 24 маркера Y-хромосомы. В качестве примера на рисунке 5 представлена медианная сеть линий 12 и 16 Y-хромосомы как наиболее интересных в контексте финноугорских народов. Линия 16 практически отсутствует в западноевропейских популяциях, но частота ее высокая среди народов Балтии - эстонцев и финнов, а также у народов Волго-Уральского региона, особенно удмуртов и коми-зырян .
Рис. 5. Медианная сеть линий HG12 и HG16 Y-хромосомы, построенная для некоторых популяций Европы и Азии
Для линии 16 Y-хромосомы уровень генетического разнообразия намного выше в популяциях Восточной Европы (чуваши, татары), чем в изученных популяциях Сибири. Хотя удмурты и имеют очень высокую частоту линий 12 и 16, уровень генетического разнообразия у них невысокий по сравнению с другими популяциями Европы. Данные о низком уровне генетического разнообразия удмуртов получены и по материнской линии при анализе полиморфизма митохондриальной ДНК. Все это свидетельствует о несомненной роли эффекта основателя и дрейфа генов в демографической истории удмуртов.
Анализ распространения и разнообразия линии 16 Y-хромосомы среди восточноевропейских популяций показывает, что местом ее "рождения", возможно, является Восточно-Европейская равнина. Согласно филогеографическому анализу этой линии в Евразии, она начала распространяться с запада на восток. В то же время частота распространения линии 12 - предковой для линии 16 - меньше, чем для линии 16.
Особенно характерна для популяций Волго-Уральского региона линия 3 Y-хромосомы, частота встречаемости которой максимальна у славян (русские и поляки), а также у населения Латвии, Литвы и Эстонии [ . Таким образом, носители этой линии - этнические группы, принадлежащие к разным языковым семьям. Частота встречаемости таких групп уменьшается в направлении север (Финляндия, Швеция) - юг (Турция, Кавказ). Анализ распространения данной линии в популяциях Волго-Уральского региона подтверждает гипотезу о возможном движении населения после Ледникового периода (Last Glacial Maximum) с территории нынешней Украины, где в то время располагался один из центров потепления .
Судя по результатам анализа отцовских линий в популяциях Восточной Европы и, в частности, Волго-Уральского региона, главную роль в формировании генетического разнообразия народов, проживающих на этой территории, по-видимому, играет географическая близость, а не языковая принадлежность. И хотя многие особенности генетической близости популяций объясняются с точки зрения их географического положения, в некоторых случаях "индивидуальная" демографическая история популяции имеет существенное значение. Хороший пример - популяция удмуртов, у которых разнообразие линий Y-хромосомы и мтДНК ограничено. Принимая темпы мутирования изученных ДНК-маркеров Y-хромосомы за 2.1 х 10 -3 и длительность одного поколения за 25 лет, получаем, что наблюдаемая дисперсия гаплотипов, выявленных у современного населения Волго-Уральского региона, сформировалась приблизительно 42.5 тыс. лет назад, что соответствует времени заселения Европы человеком в эпоху верхнего палеолита.
Таким образом, исследования полиморфизма аутосомных, митохондриальных и Y-хромосомных ДНК-маркеров внесли важный вклад в понимание путей происхождения человека и рас, расселения Homo sapiens по планете, в генетическую и демографическую историю отдельных этносов и популяций. Можно надеяться, что по мере все более подробного изучения свойств конкретных маркеров ДНК будут появляться дополнительные возможности для изучения генетической истории народов Европы и Азии. Дальнейшее развитие этногеномики в сочетании с палео- и археогеномикой значительно расширит наши представления о генофонде человека, внесет весомый вклад в понимание вопросов исторического развития и эволюции человечества.
ЛИТЕРАТУРА
1. Rosser Z.H., Zerjal Т., Hurles M.E. et al. Y-chromosomal diversity in Europe is clinal and influenced primarily by geography, rather than by language // Am. J. Hum. Genet. 2000. V. 67. P. 1526-1543.
2. Thomson R., Pritchard J., Shen P., Oefner P., Feldman W. Recent common ancestry of human Y chromosomes: Evidence from DNA sequence data // Proceedings of National Academy of Sciences. 2000. V. 97. №13 P. 7360-7365.
3. Stoneking M. Progress in population genetics and human evolution//Berlin: Springer, 1997. P. 164.
4. Cavalh-Sforza L.L. Genes, Peoples, and Languages. N.Y.: North Point Press, 2000.
5. Cruciani F., Santolamazza P.. Shen P. et al. A back migration from Asia to Sub-Saharan Africa is Supported by high- resolution analysis of Human Y-chromosome hap-lotypes //Am. J. Hum. Gen. 2002. V. 70. P. 1197-1214.
6. Лимборская C.A., Хуснутдинова Э.К.. Балановская Е.В. Этногеномика и геногеография народов Восточной Европы. M.: Наука, 2002.
7. Macualy V.A., Richards M.B., Forster P. et al. The Emerging Tree of West Eurasian mtDNAs: A Synthesis of Control Region Sequences and RFLPS //Am. J. Hum. Genet. 1999. V. 64. P. 232-249.
8. Wallace D.C., Brown M.D., Lott M.T. Mitochondrial DNA variation in human evolution and disease // Gene. 1999. V. 238. P. 211-230.
9. Бермишева M., Тамбетс К., Виллемс P., Хуснутдинова Э. Разнообразие гаплогрупп митохондриальной ДНК у народов Волго-Уральского региона // Молекулярная биология. 2002. № 6. С. 990-1001.
10. Villems R., Rootsi S., Khusnutdinova E. el al. Archaeo-genetics of Finno-Ugric speaking populations // The Roots of Peoples and Languages of Northern Eurasia. IV. Ed. by K. Julku. Oulu. 2002. P. 271-284.